18 апрель 2024
"Акушердик " адистиги боюнча иш-чара болуп өттү.
Кыргыз Республикасынын Өкмөтүнүн 2019-2030- жылдарга калктын саламаттыгын сактоо системасын өнүктүрүү боюнча "Дени сак адам - өнүккөн өлкө" программасынын алкагында медайымдык жана акушердик билим берүүнү жакшыртуу максатында2024-жылдын 17- апрелинде Бишк

15 апрель 2024
Жамбыл облусунун 85 жылдыгы
2024-жылдын 12-апрелинде боордош Казак Республикасынын Жамбыл облусунун 85 жылдыгынын урматына медициналык колледждер арасында студенттердин эл аралык “Мыкты медайым-2024” аталышындагы конкурсу болуп өттү. Иш-чараг

15 апрель 2024
Эл аралык кызматташуу, өнүгүүгө өбөлгө
2024-жылдын 11-апрелинде Акматкан Молдогазиев атындагы Бишкек медициналык колледжинин "Лабораториялык диагностика" бөлүмүнүн 3 курсунун студенткасы Султанова Амина Талдыкорган жогорку медициналык колледжинде (Республика Казахстан) өткөн "Лабор

6 апрель 2024
Дени сак адам- өнүккөн өлкө.
Дени сак адам- өнүккөн өлкө.Бүгүн, 2024-жылдын 6- апрелинде Бишкек шаарында жайгашкан Газпром спорт комплексинде, бүткүл дүйнөлүк «Ден соолук» күнүнө арналган Кыргыз Республикасынын Саламаттык сактоо министрлиги тарабынан уюштурулган «Менин ден-соолугум-
29 март 2024
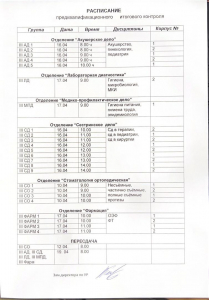
Расписание предквалификационного контроля
Расписание

29 март 2024
Международной научно-практическая конференция "МИРРАХИМОВСКИЕ ЧТЕНИЯ"
Преподаватели ПЦК»Терапия» участвовали на Международной научно-практической конференции. «МИРРАХИМОВСКИЕ ЧТЕНИЯ»Международная медицинская конференция стран ШОСС 27.03-29.03.2024г.Преподаватели ПЦК»Терапия»- Модокулова С. А, Мадымарова М. Ж., Буларкиева Г.

29 март 2024
Биринчи байлык ден -соолук!!!
Бүгүн, 2024-жылдын 29-мартында Акматкан Молдогазиев атындагы Бишкек медициналык колледжинин жыйындар залында, окуу жайдын баардык бөлүмдөрүнүн 1,2- курстарынын студенттери, окутуучулары жана бөлүм башчылары менен Кыргызстандын кызыл ай коомунун саламаттык

28 март 2024
Круглый стол на тему: "Психологические детерминанты мотивации в контексте учебной деятельности студентов – медиков"
27. 03. 2024года состоялось заседание Круглого стола в онлайн – формате на тему: «Психологические детерминанты мотивации в контексте учебной деятельности студентов – медиков». Организатором круглого стола был Уральский медицинский колледж (г.Челябинск, Ро